
2023年9月16日,3200威尼斯vip姜伊娜研究员课题组和中国科学院分子植物科学卓越创新中心王二涛研究员课题组合作在Nature Communications 杂志在线发表题为Control of arbuscule development by a transcriptional negative feedback loop in Medicago的研究论文。该研究发现,AP2/ERF蛋白ERM1/WRI5a-ERF12-TOPLESS转录复合体形成一个自我调节的正-负反馈环,动态调控营养交换和丛枝发育过程,从而维持互惠共生关系的稳定。

植物-丛枝菌根真菌共生是自然界中保守和广泛存在的共生形式。碳源与磷营养的交换是植物-丛枝菌根共生的核心。2017年,《Science》杂志的研究成果,首次揭示了丛枝菌根真菌与植物共生过程中,脂肪酸是植物传递给菌根真菌的主要碳源形式,推翻了教科书中的“糖”理论,姜伊娜为第一作者(Jiang et al., 2017, Science)。在随后的工作中,发现AP2/ERF超家族转录因子WRI5a通过靶向植物脂肪酸转运蛋白STR和磷酸盐转运蛋白PT4编码基因启动子中的AW-box元件来同时激活其表达,从而作为启动丛枝菌根共生双向营养交换的分子开关(Jiang et al., 2018, Mol. Plant)。然而,植物脂肪酸的合成、代谢及输出是一个耗能的过程,在丛枝由分枝、成熟到退化的连续发育阶段中,植物必须根据自身营养状态和需求,从时空和强度上对营养交换的速率进行动态调节,从而维持互惠共生的稳定。
本研究首先证明Half-size ABCG转运蛋白STR通过与STR2形成转运二聚体,介导苜蓿脂肪酸向丛枝细胞供应。接下来,以STR2在丛枝菌根真菌共生中的功能及其调控机制为起点展开研究,鉴定到ERF家族两个新的转录因子ERM1和ERF12拮抗调控营养交换基因表达和丛枝发育。其中,ERM1通过靶向AW-box与AW-box-like元件激活下游脂质合成及转运相关的靶基因表达,正调控菌根共生。ERF12是植物中鉴定到的首个负调控丛枝营养交换的转录因子,可通过招募TOPLESS家族辅抑制因子TPR3a,负调控菌根共生。
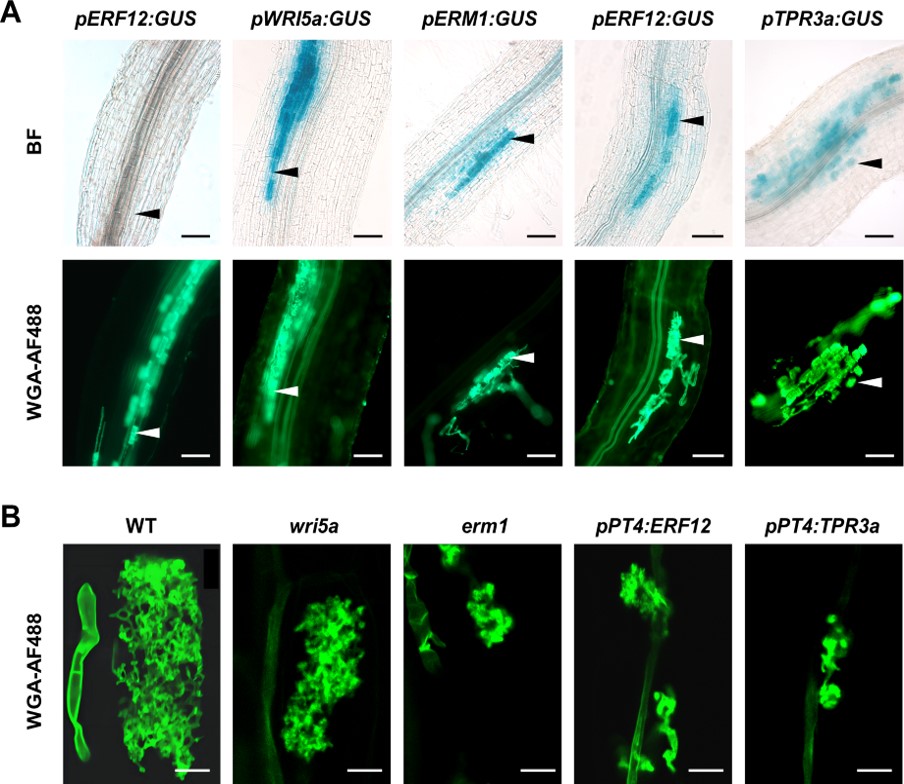
图一. 苜蓿中参与共生营养交换的关键调控因子鉴定
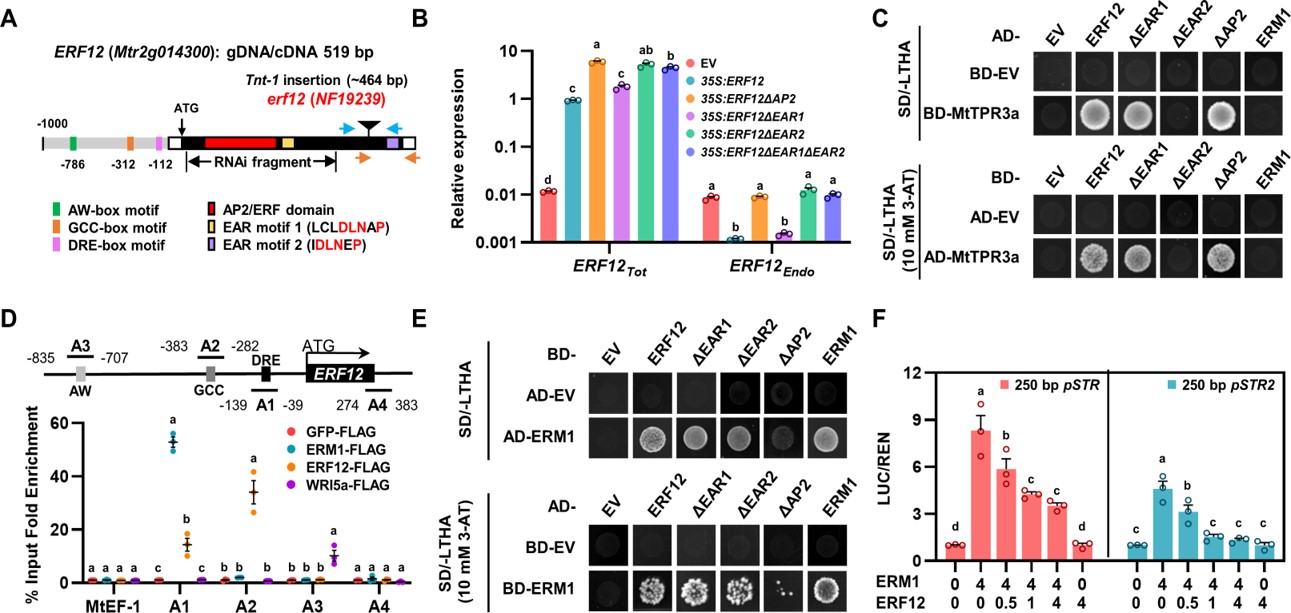
图二. ERM1/WRI5a–ERF12–TOPLESS复合体调控机制解析
接下来,作者探究了ERF12与ERM1拮抗调控丛枝菌根真菌共生的分子机制。实验结果表明,AP2/ERF蛋白ERM1和WRI5a在菌根共生早期的营养交换中发挥正调控作用,并在共生后期被重新分配以避免自身碳源的过度输出:ERM1和WRI5a在共生后期适度激活ERF12的表达,ERF12作为桥梁蛋白,提供了ERM1/WRI5a与TOPLESS家族辅抑制因子的直接联系,从而对脂质转运过程“踩下刹车”,防止植物自身资源的无效输出。此外,ERF12作为一个自我调节的转录因子,通过其AP2-DNA结合域和C端EAR基序介导对自身的转录抑制,以避免其对菌根共生进行持续或不恰当的抑制。这种“油门-刹车”反复进行的动态调控模式,初步回答了植物在共生启动后如何保护自身营养不会过度输出的关键科学问题。
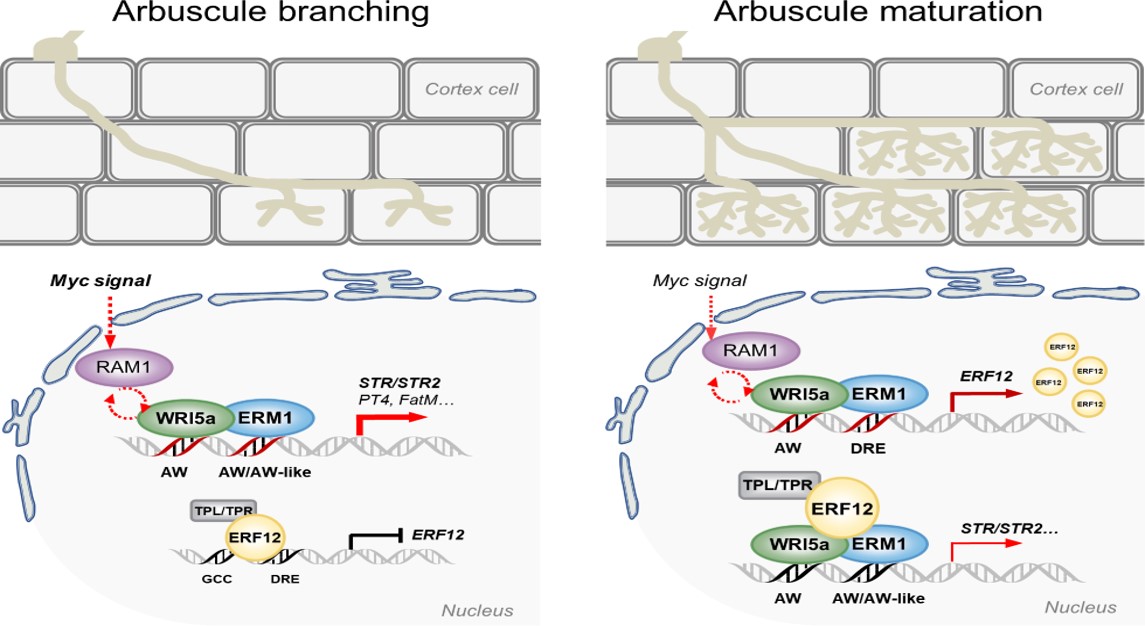
图三. 共生过程中ERM1/WRI5a–ERF12–TOPLESS复合体动态调控营养转运模型
近年来,“脂肪酸理论”在菌根共生研究领域中已获得广泛共识,一些新版和在编教材以及该领域的综述文章,已经对这一核心问题进行了修正。本研究成果的发表,加深了我们对于菌根共生中植物营养交换调控机制的理解,并为菌根共生的遗传改良及农业实践提供了重要的理论依据。3200威尼斯vip张强博士、王双双副研究员和中国科学院大学谢秋瑾博士为本论文共同第一作者,3200威尼斯vip姜伊娜研究员和中国科学院分子植物科学卓越创新中心王二涛研究员为本文的共同通讯作者,3200威尼斯vip许玲教授参与了该课题。本研究得到了国家自然科学基金等相关项目的支持。
原文链接:
https://www.nature.com/articles/s41467-023-41493-2